Apa perbedaan antara jalur alternatif klasik dan lektin
Itu perbedaan utama Antara alternatif klasik dan jalur lektin adalah bahwa inisiasi jalur klasik terjadi melalui pengikatan kompleks antibodi antigen dengan protein C1Q, dan inisiasi jalur alternatif terjadi melalui pengikatan C3B ke permukaan asing, sementara inisiasi jalur lektin terjadi melalui pengikat mannose mannose terjadi pada mannose ke mannose ke mannose ke permukaan asing, sementara inisiasi lektin terjadi melalui mannose binding mannose mannose lektin.
Jalur komplemen atau kaskade komplemen adalah bagian dari sistem kekebalan tubuh yang meningkatkan kemampuan sel dan antibodi fagositik untuk menghancurkan dan membersihkan mikroba dan sel -sel yang rusak dari suatu organisme, meningkatkan peradangan, dan menyerang membran sel patogen. Jalur komplemen dihasilkan oleh sistem kekebalan tubuh bawaan dan adaptif. Sistem ini terdiri dari protein kecil yang disintesis oleh hati dan bersirkulasi dalam darah dalam bentuk yang tidak aktif. Protein atau prekursor ini diaktifkan di jalur komplemen. Ada tiga jenis jalur komplemen: jalur klasik, jalur alternatif, dan jalur lektin.
ISI
1. Ikhtisar dan Perbedaan Utama
2. Apa jalur klasik
3. Apa jalur alternatif
4. Apa itu jalur lektin
5. Kesamaan - alternatif klasik dan jalur lektin
6. Jalur lektin vs alternatif vs alternatif dalam bentuk tabel
7. Ringkasan - jalur klasik vs alternatif vs lektin
Apa jalur klasik?
Jalur klasik adalah salah satu dari tiga jalur yang mengaktifkan sistem komplemen. Sistem komplemen adalah bagian dari sistem kekebalan tubuh. Kompleks antigen-antibodi bersama dengan isotipe antibodi IgG dan IgM memulai sistem komplemen. Sel apoptosis, sel nekrotik, dan protein fase akut juga mengaktifkan jalur klasik.
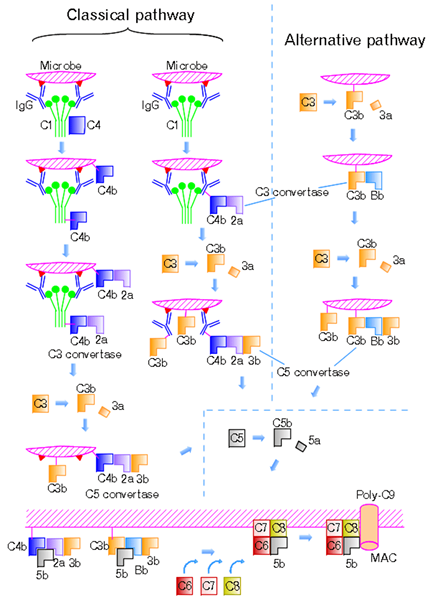
Gambar 01: Jalur klasik dan jalur alternatif
Jalur ini diprakarsai oleh pengikatan kompleks antigen-antibodi dengan protein C1Q; Wilayah globular C1Q mengenali dan mengikat ke wilayah FC dari antibodi IgG dan IgM isotipe. Mereka juga mengikat protein permukaan bakteri dan virus, sel apoptosis, dan protein fase akut. Selama tidak adanya faktor aktivasi, C1Q menjadi bagian dari kompleks C1 yang tidak aktif, yang terdiri dari enam molekul C1Q, dua molekul C1R, dan dua molekul C1s. Ikatan C1Q menyebabkan perubahan konformasi dan aktivasi serin protease C1R. Ini mengaktifkan dan memecah serin protease c1s. C1 kemudian membelah C4 menjadi C4A dan C4B dan C2 menjadi C2A dan C2B. Aids C4B dalam pembentukan C3 convertase, C4BC2A. Konversi C3 memiliki kemampuan untuk membelah C3 menjadi C3A dan C3B, yang merupakan faktor penting untuk reaksi enzimatik berikutnya. C3V berikatan dengan C3 convertase untuk membentuk C5 convertase, C4B2A3B, sementara C3A merekrut sel -sel inflamasi melalui. Ini dikenal sebagai anaflatoksin. C5 convertase membelah C5 menjadi C5 A dan C5B. C5B bergabung dengan komponen terminal lainnya untuk membentuk kompleks serangan membran (MAC). Hal ini menyebabkan lisis bakteri invasif dengan memasukkan ke dalam membran sel target, menciptakan pori -pori fungsional.
Apa jalur alternatif?
Jalur alternatifnya adalah salah satu dari tiga jalur yang menyingkirkan dan menghancurkan patogen. Virus, jamur, bakteri, parasit, imunoglobulin A dan polisakarida mengaktifkan jalur alternatif dan membentuk mekanisme pertahanan penting yang terlepas dari sistem kekebalan tubuh. Protein C3B memicu jalur ini, dan protein ini secara langsung berikatan dengan mikroba. Bahan asing dan jaringan yang rusak juga memicu jalur alternatif. Karena C3B bebas dan berlimpah dalam plasma, ia memiliki kemampuan untuk mengikat sel inang atau permukaan patogen. Protein pengatur yang berbeda mengambil bagian dalam pencegahan aktivasi komplemen pada sel inang.
Reseptor komplemen 1 (CR1) dan faktor pelengkap-akselerasi (DAF) bersaing dengan Faktor B untuk mengikat dengan C3B pada permukaan sel dan menghilangkan BB dari kompleks C3BBB. Pembelahan C3b ke dalam bentuk tidak aktif, IC3B, dengan protease plasma yang disebut faktor komplemen 1 menghambat pembentukan c3 convertase. Faktor komplemen 1 membutuhkan kofaktor protein pengikat C3B seperti faktor H, CR1, atau kofaktor membran proteolisis. Faktor H menghambat pembentukan C3 convertase dengan bersaing dengan Faktor B untuk mengikat dengan C3B. Ini juga mempercepat pembusukan convertase C3. CFHR5, yang merupakan faktor komplemen H terkait protein 5, memiliki kemampuan untuk berikatan untuk bertindak sebagai kofaktor untuk faktor 1 dan mempercepat aktivitas yang membusuk, dan berikatan dengan C3B pada sel inang.
Apa itu jalur lektin?
Jalur lektin adalah jenis reaksi kaskade dalam sistem komplemen. Setelah aktivasi jalur ini, aksi C4 dan C2 menghasilkan protein komplemen yang diaktifkan lebih jauh ke bawah kaskade. Jalur ini tidak mengenali antibodi yang terikat pada targetnya dan dimulai dengan lektin pengikat mannose (MBL) atau ficolin yang mengikat gula tertentu. MBL ini berikatan dengan gula seperti mannose dan glukosa dengan kelompok OH dalam posisi terminal pada karbohidrat atau komponen glikoprotein bakteri, jamur, dan beberapa virus.
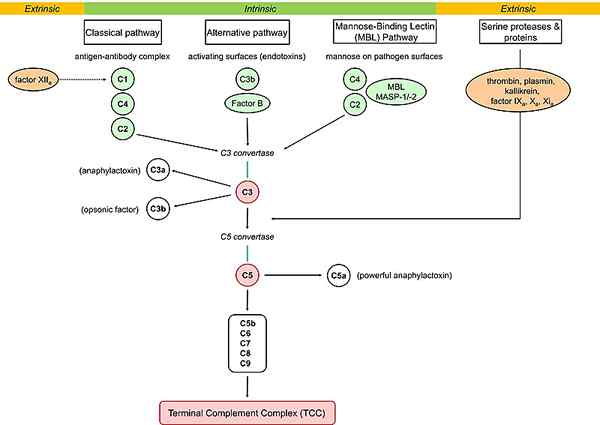
Gambar 02: Jalur komplemen
MBL, juga dikenal sebagai protein pengikat mannose, mampu memulai sistem komplemen dengan mengikat permukaan patogen. Multimer MBL membentuk kompleks dengan protease serin (mannose pengikat lektin terkait serin protease: masp1, masp2 dan masp3) yang merupakan zymogen protein. Mereka mirip dengan C1R dan C1 di jalur lain. MASP1 dan MASP2 Aktifkan untuk membelah komponen C4 dan C2 menjadi C4A, C4B, C2A, dan C2B. C4B cenderung mengikat membran sel bakteri. Jika tidak diaktifkan, ia bergabung dengan C2A untuk membentuk convertase C3 convertase convertase klasik convertase alternatif. C4a dan C2B bertindak sebagai sitokin yang kuat. C4a menyebabkan degranulasi sel mast dan basofil, dan C2B meningkatkan permeabilitas pembuluh darah.
Apa kesamaan antara alternatif klasik dan jalur lektin?
- Jalur klasik, alternatif, dan lektin diaktifkan oleh kaskade reaksi yang mengarah ke kompleks serangan membran.
- Mereka adalah bagian dari sistem kekebalan tubuh.
- Setiap jalur memiliki protein unik untuk inisiasi.
- Mereka diaktifkan oleh isotipe antibodi tertentu yang terikat pada antigen
Apa perbedaan antara jalur alternatif klasik dan lektin?
Inisiasi jalur klasik terjadi melalui pengikatan kompleks antibodi antigen dengan protein C1Q. Inisiasi jalur alternatif terjadi melalui pengikatan C3B ke permukaan asing, sementara inisiasi jalur lektin terjadi melalui lektin yang mengikat mannose. Dengan demikian, ini adalah perbedaan utama antara alternatif klasik dan jalur lektin. Peran jalur klasik adalah bahwa ia bertindak sebagai lengan efektor kekebalan adaptif sementara jalur alternatif dan lektin berfungsi dalam kekebalan bawaan. Selain itu, aktivasi C4 dan C2 di jalur klasik adalah C1s, dan di jalur lektin adalah MASP-2, sedangkan tidak ada aktivasi C4 dan C2 di jalur alternatif.
Infografis di bawah ini menyajikan perbedaan antara jalur alternatif klasik dan lektin dalam bentuk tabel untuk perbandingan berdampingan.
Ringkasan -jalur klasik vs alternatif vs lektin
Inisiasi jalur klasik terjadi melalui pengikatan kompleks antibodi antigen dengan protein C1Q. Inisiasi jalur alternatif terjadi melalui pengikatan C3B ke permukaan asing sementara inisiasi jalur lektin terjadi melalui lektin pengikat mannose. Jadi, ini adalah perbedaan utama antara alternatif klasik dan jalur lektin.
Referensi:
1. “Jalur Klasik.“Biolab kreatif.
2. Fujita, Teizo. “Evolusi jalur pelengkap lektin dan perannya dalam kekebalan bawaan.Berita Alam, Grup Penerbitan Alam.
Gambar milik:
1. “Jalur pelengkap.”Oleh Tossh_eng - https: // unggah.Wikimedia.org/wikipedia/commons/b/b0/jalur pelengkap.PNG (CC BY-SA 4.0) Via Commons Wikimedia
2. "Gambar 1.4. Aktivasi komplemen ”oleh Hofman.Martijn - Karya Sendiri (CC BY -SA 4.0) Via Commons Wikimedia